糖鎖は、各種の単糖が文字通り鎖のようにつながった糖質の一つで、生体内でタンパク質や脂質と結合することでそれらの機能を調整するとともに、細胞認識や細胞接着といった生命維持に関わるプロセスでも重要な役割を果たす。タンパク質の代謝時期を決める機能も有していて、体内のタンパク質は結合する糖鎖によって生体内や血中内での寿命が決まる。一方、生物に多様な進化をもたらした核酸、タンパク質に次ぐ、生命現象の謎を解く第3の「生命鎖」としても注目される。
特集 「糖質」そこが知りたい 代謝で重要な役割を担う「糖鎖」 生命現象を紐解く鍵となるか
構成/飯塚りえ イラストレーション/小湊好治

東洋大学食環境科学部食環境科学科教授
宮西伸光(みやにし・のぶみつ)
1999年、長崎大学大学院水産学研究科修了。2003年、オーストラリア・フリンダース大学研究員、同年、東京水産大学大学院博士課程修了および香川大学総合生命科学研究センター教員。2008年、同センター客員准教授。2010年から東洋大学生命科学部食環境科学科(現・食環境科学部食環境科学科)准教授、2015年から現職。2018年、英国インペリアル・カレッジ・ロンドン研究員。2021年、 横浜市立大学客員教授併任。
糖鎖は、グルコースやマンノースなどの単糖類が数個から数十個連なり、通常はタンパク質や脂質に結合した形で存在して、細胞表面を覆っています。
生体のチューニングという重要な役割を担う糖鎖は、生体内や細胞内で起こったイベントに対処すべく生物が作り出したツールと考えられます。その身近な例が血液型です。ABO式の血液型は、その人の持っている糖鎖の種類によって決まります。これは遠い祖先の感染の痕跡といわれています(図1)。
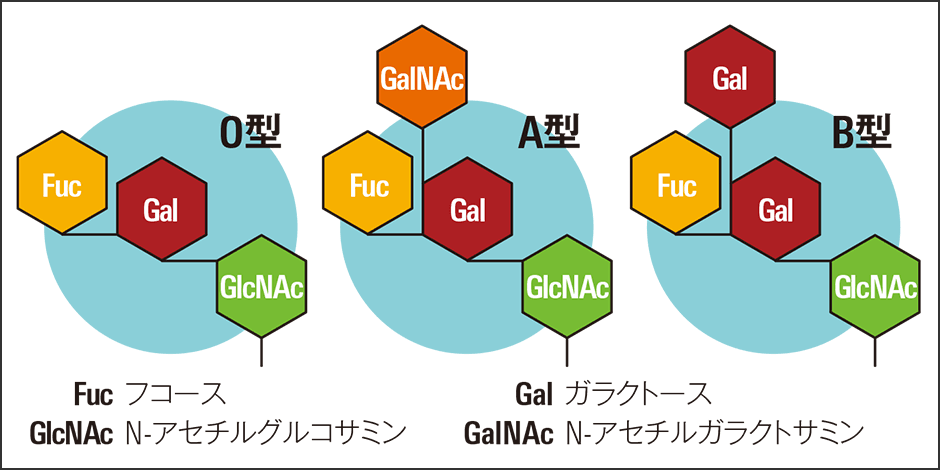
図1 血液型血液型は赤血球の表面にくっついている糖鎖によって決まる。O型が持つ糖鎖に、何らかの感染に対処するためN-アセチルガラクトサミン(GalNAc)がくっついたと考えられているものがA型、同様の経緯でO型の糖鎖にガラクトース(Gal)がもう一つくっついたものがB型。AB型はこの両方の糖鎖を持っている。
タンパク質や脂質の機能を調整
糖鎖の多岐にわたる役割を見ていきましょう。
糖鎖は、細胞認識や細胞接着といった生理的な機能に関わるほか、結合しているタンパク質や脂質の機能の調整を担います(図2)。
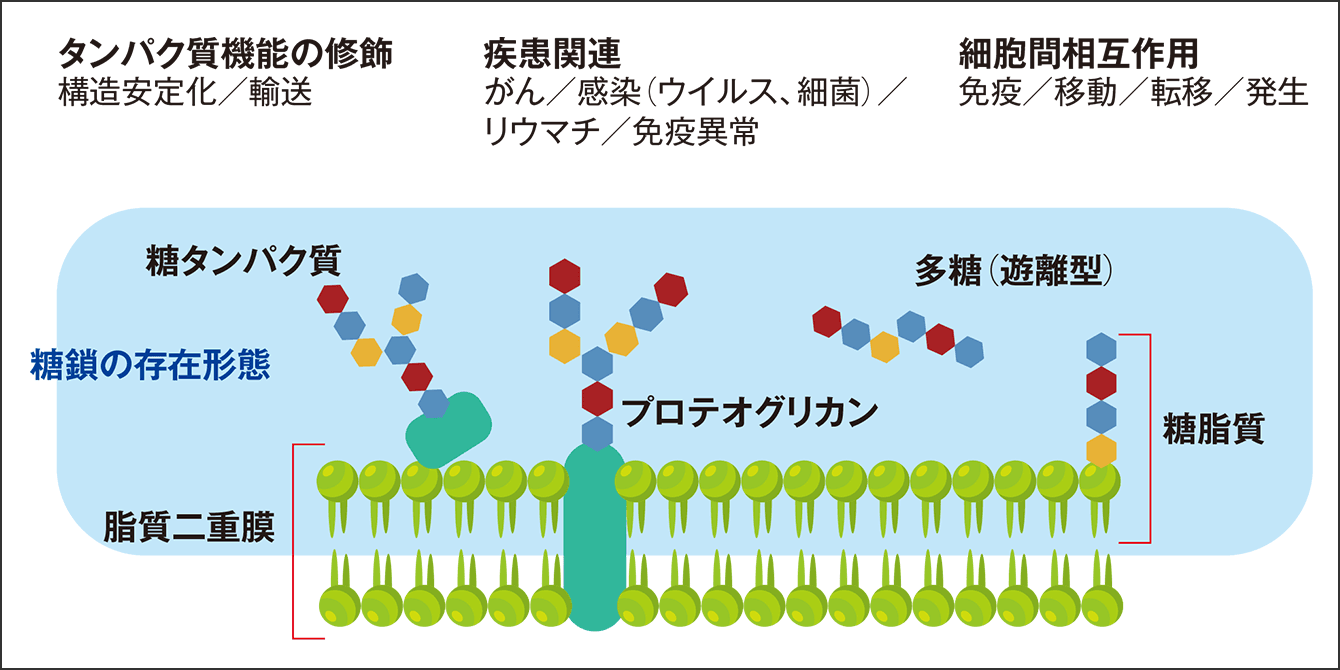
図2 細胞社会における多様な糖鎖の機能糖鎖は、細胞の膜タンパク質に付いていたり、抗体の中に入っていたりと、タンパク質や脂質にほぼ結合した形で存在し、タンパク質機能の修飾、疾患、細胞間の相互作用といった生体内でのチューニングを担う。
まず、糖鎖には、タンパク質機能を修飾して細胞の行き先を明確にするという機能があります。細胞内で作られた多様なタンパク質は、それぞれ糖鎖に修飾されて、あるべき部位まで運ばれます。タンパク質の機能を示す目印の役割を果たしているのです。
糖鎖はまた、タンパク質の構造安定化という物理的な支えも担います。細胞の構造がしっかりと維持されていないとその細胞は機能を発揮することができません。
また、タンパク質の代謝時期を決めるという機能もあります。体内のタンパク質は、修飾される糖鎖によって生体内や血中内での寿命が決まります。結合する糖鎖が1個外れただけで、すぐに代謝されて排出されたり、逆に数日だった寿命が1週間以上延びたりといったことが起こります。
疾患関連では、がん、リウマチ、感染、免疫異常といった自己と非自己を認識する目印として糖鎖が利用されています。体内のチューニングという“揺らぎのある現象”を担う糖鎖が、自己/非自己の認識という、境界線の曖昧な仕事に関わるのは当然といえます。
感染の場合、原因となるウイルスや菌は細胞表面を覆う糖鎖を狙います。
例えば、インフルエンザウイルスは、赤血球表面にある糖鎖の末端に結合しているシアル酸という糖を狙います(図3)。この糖鎖にくっついて細胞の中に入り、自身のRNAを増殖させて細胞外に出、発症に至ります。鳥インフルエンザウイルスの場合、α2,3型という結合方式をとる鳥の糖鎖を狙います。
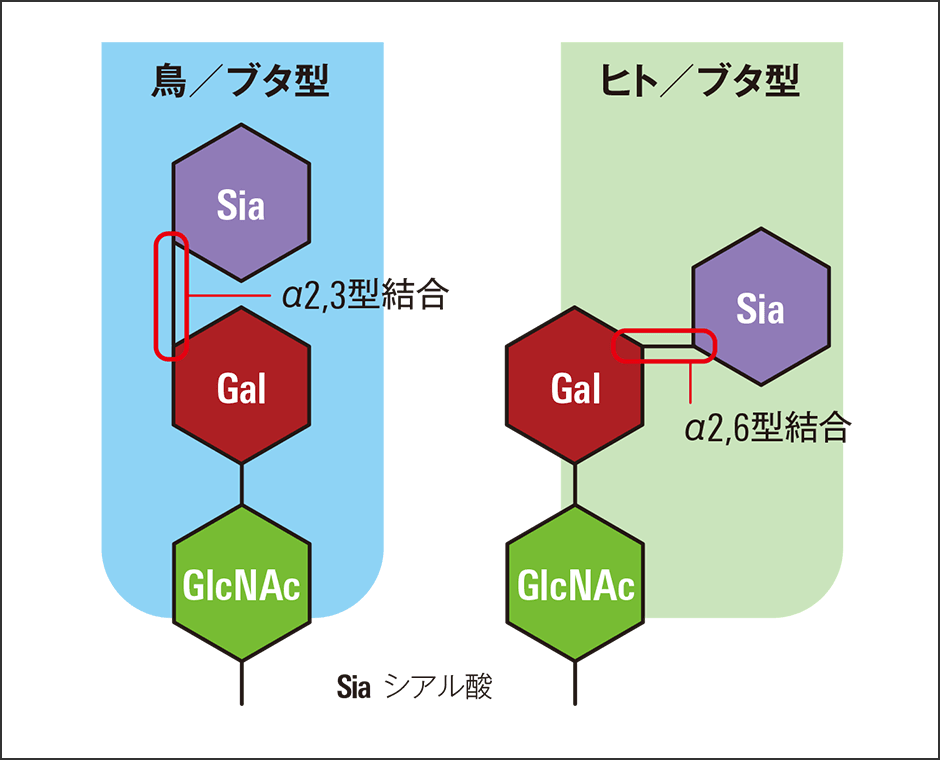
図3 インフルエンザと糖鎖の結合インフルエンザウイルスは、赤血球表面の糖鎖にあるシアル酸(Sia)をターゲットとしている。糖鎖は感染においても重要な役を演じている。
進化を支えるために作り出された
一方、ヒトの糖鎖はα2,6型の結合なので、このままでは鳥インフルエンザウイルスがヒトに感染することはできません。しかし、やっかいなことに、ブタの糖鎖にはα2,3型、α2,6型の両方の結合があります。そのため、鳥インフルエンザウイルスはブタに感染することが可能で、さらにブタの体内で変異を起こせば、鳥インフルエンザウイルスがヒトに感染できるようにもなります。このウイルスが強毒化すれば、新型コロナウイルス感染症同様のパンデミックが起きる可能性があるのです。
ではなぜ、糖鎖はこのように多様な機能を担うことになったのでしょうか。
糖鎖は、外界や細胞内外とのインタラクションのために生物が作り出したものです。それはつまり、進化を支えるためだったと考えられます。
太古の地球環境において、グルコースやマンノースなどの単糖は、化学反応だけで、つまり生物の介入を必要とせずに合成することが可能だったと考えられています。ですから、生物が誕生したときにはすでに存在しており、生物はこれらの糖をエネルギー源として利用できたはずです。糖の話題になると、最初に「エネルギー源」と思い浮かぶのは、こうした長い歴史があるからでしょう。
その後、進化に伴って光合成を行う植物性プランクトンが登場し、生物は脂質を代謝してエネルギーを得るという新しいシステムを手に入れました。私たちヒトのような高度に進化した生物は、これら新旧ハイブリッドの代謝システムを使っています。
しかしそれでも、中枢神経系という生物のコアとなる領域のエネルギー源はグルコースに大きく依存しています。脂質は、エネルギー源として非常にコストパフォーマンスの高い物質だというのに、どうして利用しないのかといえば、脂質はエネルギー量が多い分、制御も難しくなるからです。
遺伝子は糖鎖によって多様性を獲得できる
先に示したように、グルコースは生物誕生以前から現在にまで存在する、際立って安定性の高い物質です。そして生物は、誕生から変わらず、常にグルコースからエネルギーを得て“無事に”進化を遂げています。だとすれば、中枢神経系という生命維持の司令塔のエネルギー源として、これほど信頼性の高い物質はないのです。太古の地球で、生物の化学反応だけで合成が可能だったのは、グルコースの他にも、マンノースやフルクトースなどがあります。これらの糖は、動植物すべてにとって、エネルギー源として重要な糖です。
しかし一方で、生物が複雑に、高度に進化するには、さまざまな“部品”が必要になります。表は種によるゲノム全体のサイズと遺伝子数を記したものです。体の大きさは、例えば大腸菌は2~4μm、ヒトは160㎝程度と約40万~80万倍ですが、ゲノムサイズは、大腸菌で4.64Mbp、ヒトは3000Mbp程度で645倍ほどです。ショウジョウバエとヒトの差でも25倍ほどです。
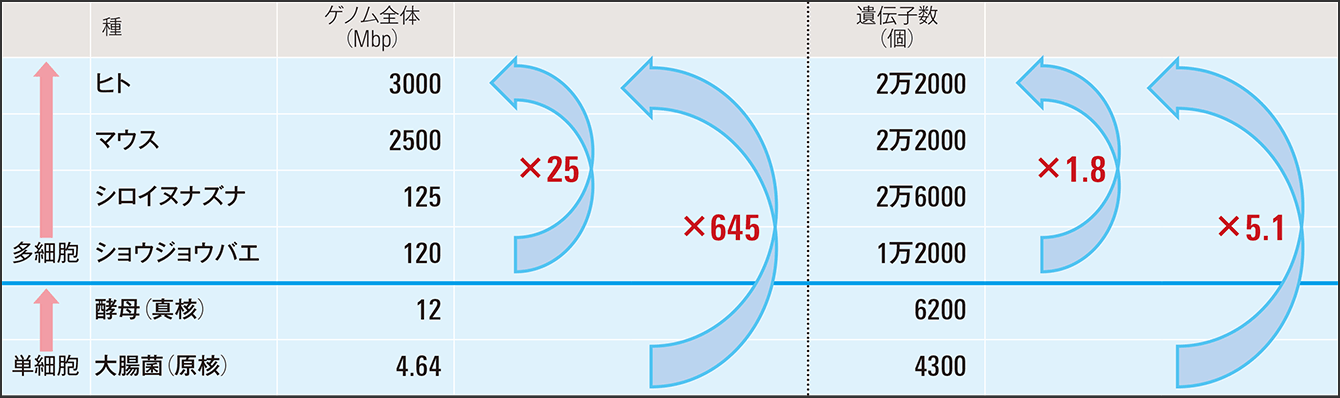
表 種によるゲノム、遺伝子数などのボリューム遺伝子や核酸だけではヒトのような複雑な種に進化するための“部品”が足りないことが分かる。
次にゲノムの中で、タンパク質がコードされている遺伝子数を比較してみます。ヒトとマウスはほとんど変わりません。ショウジョウバエとヒトの差が1.8倍、植物のシロイヌナズナになると、むしろヒトよりも多いぐらいの勢いです。
では3つの主要な生命鎖を見ましょう。核酸やアミノ酸は結合の種類が1系統しかありません。それに対して糖はOH基の結合を利用して、10系統ほどの結合を生み出すことが可能です。例えば、核酸、アミノ酸、糖がそれぞれ6個結合した場合、それぞれの結合のバリエーションは、ATGCという4種類の核酸塩基を持つDNAは4の6乗種類、アミノ酸の種類は20程度なのでペプチド(タンパク質)は20の6乗種類ですが、各種の糖を結合してできる糖鎖の種類は実に1兆以上になります。つまり、遺伝子や核酸だけでは多様性を生み出す“部品”が不足しているのです。それを十分に補ったのが糖(糖鎖)といえるでしょう。
恐らく、最初はグルコースなどの単糖を少し変化させて使っていたはずですが、進化するうちに、生物が酵素を作り出して元々の糖を改変し、変わった新しい糖を作ったり、連結させたりといったようなことができるようになりました。こうして生物の介入があって多糖類ができると、生物はそれを細胞の支持体にしたり、エネルギー貯蔵に使ったりと、糖の使い道をぐんぐんと広げていきました。その最終進化形といえるようなものが糖鎖なのです(図4)。
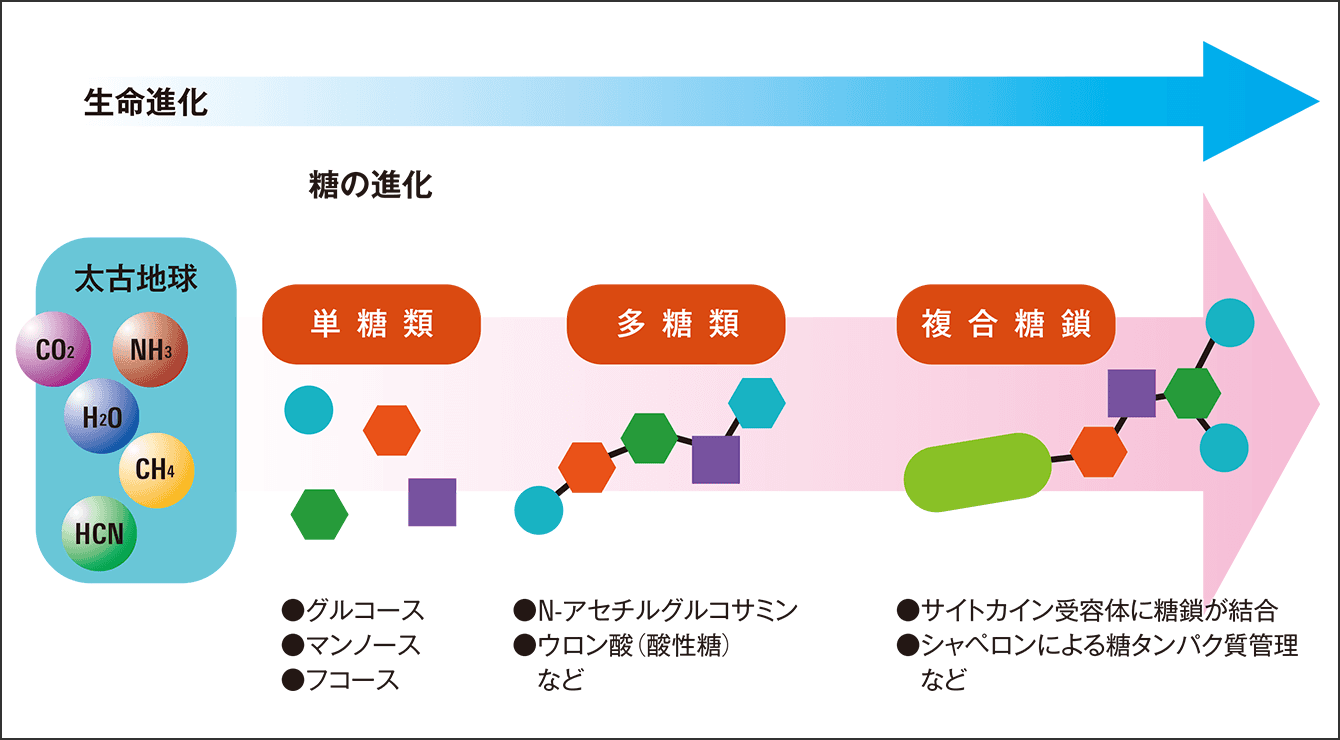
図4 生命進化に必須の糖太古地球で初期の生物は、グルコースを多少改変したN-アセチルグルコサミン(GlcNAc)など、グルコースに別の糖を結合して新しい糖を合成していったと考えられる。これで必要な“部品”が賄いきれなくなると、マンノースを使う。いっそうの多様性を求めてガラクトースやフコースなどから多糖を作り、さらに複雑な糖鎖を作っていった。ガラクトースは生物の介入なしには作れないとされており、糖と糖鎖の進化が生物の進化とリンクしていることが見える。糖鎖は、タンパク質や脂質などとともに存在することがほとんどで、さまざまな分子に結合してそれぞれの機能を調整するようになった。
糖鎖はタンパク質や脂質に結合する形で生体内に存在すると前述しましたが、アスパラギンに結合するN型糖鎖の構成糖には、実はグルコースがほぼ使われていません。それは、生物がグルコースを貴重なエネルギー源として採用しているからです。エネルギー源としての糖と、生化学的な物質としての糖や糖鎖とをしっかり区別して利用しているのですが、グルコースからさまざまな糖への進化は連綿とつながった生命の営みそのものなのです。
植物糖鎖の研究のために私たちが用いている微細藻類の赤潮プランクトンは、単細胞で運動性があり、光合成もするという動物と植物の両方の特徴を併せ持った、動植物の進化系統樹の根元に位置するような生物ですが、これらが持っているN型糖鎖には、ほぼマンノースしかありません。初期のN型糖鎖は、グルコースの改変型ともいえるN-アセチルグルコサミン(Glc NAc)とマンノースを順にくっつけていくのですが、その他の糖を付けるまでのシステムには至りません。それがコケ・シダ類に進化すると、キシロースなどのその他の糖が加わってくるようになります。
さらに、被子植物や裸子植物のように高度に進化すると、幹や果実を作るために、さまざまな情報を用いた複雑なシステムが必要になるので、多様性を生み出すための部品としてより多様な糖が用いられるようになるのです。実際、植物進化において大きなイベントがあると、コケ類からヒバ類への進化など植物のフェーズも変わります。すると同時に糖鎖も変化していることが見えてきました。
糖鎖と進化が密接に関連していることについて網羅的に解析し、実用にも生かすべく、私たちは今、穀類の糖鎖を解析しています。
イネやコムギ、トウモロコシといった主要な穀類は、収穫量を上げたり食味を変えたりするために盛んに品種改良が行われており、当然、改良型と野生型では糖鎖にも大きな違いがあります。野生型は、過酷な環境変化に適応すべく糖鎖を蓄積してきたはずですが、最大限の収穫を目的として環境を整えられた“温室育ち”の改良型穀類は、環境変化に対して脆弱になっているのです。改良型は、環境の変化を受けると、糖鎖の秩序がガタガタと崩れてしまいます。これらの糖鎖の構造解析を進めて、環境が糖鎖にどのような違いをもたらしているのかを調べています。それによって改良型の脆弱性を克服し、頑健性を取り戻すことができるのではないか、と考えています。
生物は、環境の変化に適応するために、適した糖を選んで糖鎖を作ってきました。糖鎖を理解するには、糖の進化を理解すること。それはさらに生物の進化を解明することにもつながるのです。
(図版提供:宮西伸光)