地表の約70%を占める海洋には、地球の生態系に多大な影響を与える多様な生物が生息している。中でも微小な植物プランクトンは、二酸化炭素の吸収源として大きな役割を果たしている。それは、海洋植物プランクトンが気候を左右する重要なファクターであるということにほかならない。そして植物プランクトンを制御する因子の一つが、巨大ウイルスであることはほぼ確実なのだという。巨大ウイルスと植物プランクトンの関係の解明は、今後の気候変動対策に大きく寄与するはずだ。
特集 知られざるウイルス 海洋植物プランクトンを制御する「巨大ウイルス」
構成/河﨑貴一

京都大学化学研究所バイオインフォマティクスセンター助教
遠藤 寿(えんどう・ひさし)
2008年、東京学芸大学教育学部卒業。2013年、北海道大学大学院環境科学院博士課程修了(環境科学博士)。JST-CREST博士研究員を経て、2017年から現職。
海洋は地表の約7割を占めています。その海洋には広大な生態系が存在し、水産資源の供給源や人為起源による二酸化炭素(CO2)の吸収源になっています。典型的な外洋の表層海水1㎖を調べると、真核生物が少なくとも数千~数万細胞、原核生物は数十万~数百万細胞、ウイルスは数千万~数億粒子も存在しています。
- *1 真核生物:細胞の中にDNAを包む細胞核がある生物。単細胞および多細胞の動物、藍藻類を除く植物、真核菌類など。
- *2 原核生物:核を持たない細胞(原核細胞)からなる生物。細菌と藍藻類がこれにあたる。
海洋の物質循環に影響するウイルス
海洋で最も大事な基礎生産者は微細な植物プランクトンです。近年、海洋微生物の遺伝子解析などの研究により、植物プランクトンは海洋巨大ウイルスとの相互作用を介して、大気中のCO2を固定して深海へ輸送したり、気象にも影響したりすることがわかってきました(図1)。
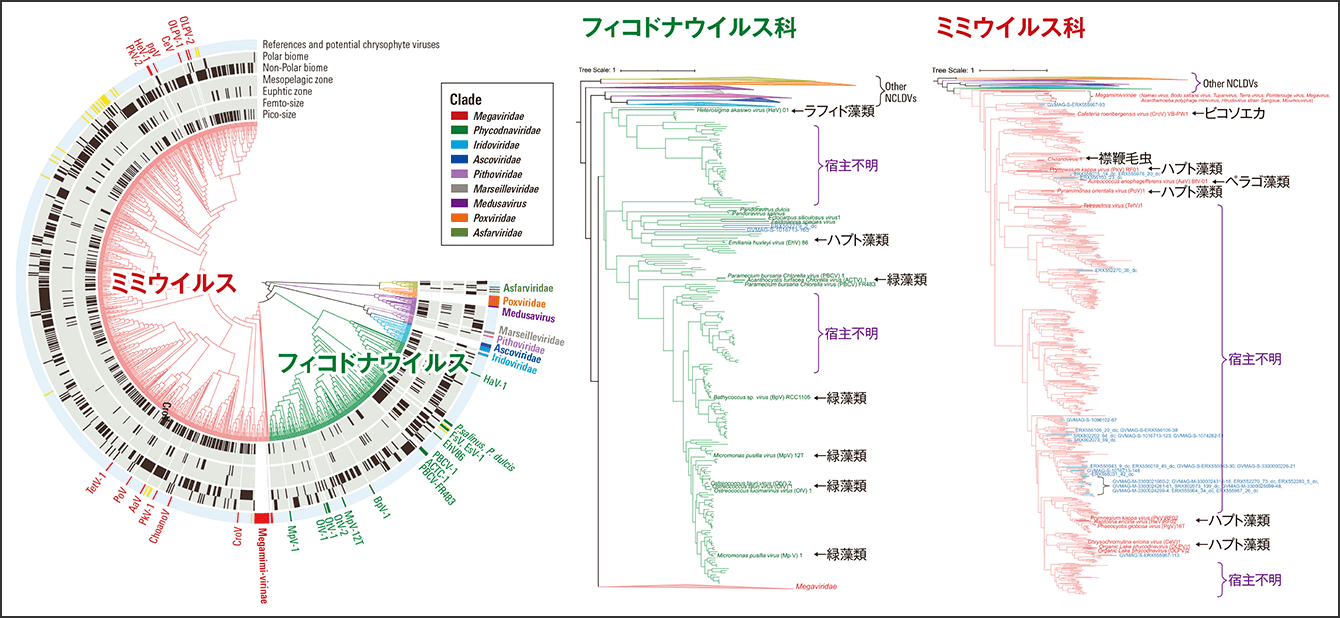 (巨大ウイルスのPolB系統樹:宿主既知系統の参照配列と905個の環境配列〈>700 アミノ酸長〉を用いて作成)
(巨大ウイルスのPolB系統樹:宿主既知系統の参照配列と905個の環境配列〈>700 アミノ酸長〉を用いて作成)
図1 海洋メタゲノムで検出された巨大ウイルス(NCLDV)の系統海洋メタゲノムの遺伝子解析による巨大ウイルスの系統樹。フィコドナウイルス科、および、ミミウイルス科(特にMesomimivirinae)の系統内で特に多様度が高い。既知宿主と遠縁の枝も多数見られ、より広い宿主系統群の存在を示唆している。
ウイルスは、生物か非生物かという議論が続いていますが、ウイルスだけではエネルギーの生産・代謝はできず、自己増殖もできません。ウイルスが増殖するためには、宿主となる生物に感染し、宿主細胞の代謝やタンパク質合成を行う分子装置を利用する必要があります。多くの場合、宿主の中で増殖したウイルスは宿主細胞を破壊して環境中に放出されます。つまり、単細胞性の微細生物にとって、ウイルス感染は死を意味するのです。海洋では、細菌から、微小な植物プランクトンや動物プランクトン、魚類に至るまで、幅広い栄養段階でウイルス感染の影響を受けています。中には、宿主の生理機能を改変することで、微生物の群集動態に影響を及ぼすものもいます。
ウイルス感染によって宿主細胞が破壊されると、細胞に含まれる有機炭素、窒素、リンなどの栄養資源は環境に放出され、従属栄養細菌の分解を介して他の生物に取り込まれたり、あるいは凝集して粒子状になって深層へ沈んでいきます。
- *3 従属栄養細菌:生育に有機炭素源を必要とする細菌。
海の深さで区分すると、表層(0~200m)、中深層(200~1000m)、それ以下に大きく分けられます。表層の水深50~150mぐらいまでは太陽光が届く有光層で、ここでは植物プランクトンが大気から海水中に溶解したCO2を使って光合成を行います。植物プランクトンは、光合成によってCO2からグルコース(C6H12O6)などの有機物を生成します。また、一部の植物プランクトンはCO2が解離した炭酸イオン(CO32-)と海水中のカルシウムイオン(Ca2+)から炭酸カルシウム(CaCO3)を作ります。この反応の際に、炭酸系の平衡反応によってCO2が生成されるため、石灰化は光合成と逆にCO2を排出する反応ともいえます。つまり、海洋表層に存在する植物プランクトンの種や生物量を調べると、その海域の炭素循環の性質を推定することができます。
海中の遺伝学的データを解析する
私たちが研究しているのは海洋中の「巨大ウイルス」と呼ばれるウイルスで、比較的大きな粒子やゲノムを持っています。巨大ウイルスは正式なウイルス分類群の名称ではなく、それを定義する上でのゲノムや粒子サイズによる明確な境界線はありませんし、外見上の特徴も種によってさまざまです。一般的に巨大ウイルスと呼ばれるものは、粒子サイズが200nm以上、ゲノムサイズは100kbp以上のものがほとんどですが、それより小さなウイルスも含まれます。
- *4 bp:塩基対(base pair)。
すべての巨大ウイルスはヌクレオサイトウイルス(Nucleocytoviricota)門に属するとされています。しかし、近年あらゆる環境から巨大ウイルスの新種が続々と分離されており、国際ウイルス分類委員会(ICTV)による正式学名の付与を待たずに実質的に巨大ウイルスとみなされているウイルスも数多く存在します。
巨大ウイルスは、2003年、フランスのエクス=マルセイユ大学の微生物学者、ベルナール・ラ・スコラらによって発表されたミミウイルス(Acanthamoeba polyphaga mimivirus)が最初でした。実は、ミミウイルスは1992年に発見されていましたが、光学顕微鏡でも観察できるほど大きかったので、細菌と考えられていました。
その後、次世代シーケンサーが開発されて、ゲノム解読が急速に進歩しました。生物のゲノムは通常、数百万以上もの長い塩基で構成されていて、それを読むためには大量の配列を読み取る必要がありました。現在稼働している次世代シーケンサーは数十億配列もの情報を一度に取得できるため、生物のゲノム配列を決定するのに十分な情報が安価に得られる時代になっています。ただし、現在主流となっているショットガン・シーケンス法では、制限酵素や超音波で短くしたDNA配列を読むため、コンピュータでつなぎ合わせて元の配列を復元する作業が必要です。いわば、シュレッダーでバラバラになった紙を組み合わせて、元の文章に復元する作業に似ています。
次世代シーケンサーを使うことで、数万種類といわれる海洋プランクトンを分類したり、微小なプランクトンを分離することなく、微生物の遺伝子を早く、かつ網羅的に解読できるようになりました。そして、微生物やウイルス群集におけるデータの客観性や定量性を、ある程度担保できるようになりました。まだ課題もありますが、観測情報のデータベース化が進んでいます。
近年では、さらに進化したロングリード・シーケンサーが開発されて、長いDNA配列を断片化せずに、そのまま解読できるようになりました。このロングリード・シーケンサーは、今後の遺伝子の研究には主流になると思いますし、私も海洋生態系研究で利用しています。
海洋生態系の重要な機能
巨大ウイルスの研究については、Tara Oceans(タラ海洋探査)と呼ばれる国際的な海洋探査プロジェクトの一環として、2009年から2013年にかけて海洋の大部分を網羅する調査航海が行われました。調査の主役になったのは、海水中のDNAやRNAなどの遺伝学的データです。調査地点は南極海から北極海までの120地点を数えます。
当時、北海道大学の大学院生だった私は、たまたま国際学会で居合わせたプロジェクト・コーディネーターから航海に誘われましたが、そのときは他の研究もあって参加できませんでした。ところが、その6年後に現在の京都大学化学研究所に移ってから、このプロジェクトのデータ解析を担当することになったのは、何かの縁だったと思います。

南極海で試料採取中の遠藤助教(左)。
Tara Oceansの海洋調査プロジェクトで採取された約300の微生物群集に由来するDNAを、次世代シーケンサーで半網羅的に読み取ったのがメタゲノムデータです。現在では、Tara Oceans海洋探査プロジェクトで採取した370の海洋メタゲノムデータから、約4700万の遺伝子配列に集約した海洋遺伝子のカタログが構築されています。私たちは、この遺伝子カタログの中から巨大ウイルスの指標遺伝子であるDNAポリメラーゼBの配列を探索し、未知の種を多数含む約7000種(系統型)の巨大ウイルスを検出しました(図1)。
- *5 DNAポリメラーゼB:一本鎖DNAを鋳型として二本鎖DNAを合成する酵素の一つ。
海洋は、表層の50~100mぐらいは海水が混ざっていますが、それ以下はほとんど混ざりません。表層から中深層に物質が移動するためには、重力によって自重で沈降するか、魚類などの大型生物の鉛直(垂直)移動に乗って運ばれる必要があります。
ところが、私たちの解析データを見ると、表層と中深層で、同じようなウイルスが出てくる観測点がたくさんありました。どうやら、ウイルスは上から下に移動しているようですが、比較的サイズが大きい巨大ウイルスでも、自重で下に落ちるほど重くはありません。巨大ウイルスが下に移動するには、もっと大きく重い宿主に感染する必要があります。例えば、プランクトンなどの凝集体が下層に炭素化合物を運ぶ生物炭素ポンプに関与していると考えるのが妥当です。
ウイルス感染によってストレスを受けたり、死滅したプランクトンは凝集体を形成する性質があります。海洋表層で光合成によって増えたプランクトンは死滅・凝集して沈んでいくことにより、大気中から海水に溶解したCO2を、数百年から数千年の期間、深層へ隔離します。この機能は生物炭素ポンプと呼ばれます。この働きがなければ大気中のCO2は現在の倍の濃度になっているとも試算され、海洋生態系は気候を安定的に維持する重要な機能を果たしています。
植物プランクトンの制御因子
カナダの研究者は、ウイルスが宿主の死滅と凝集を促進させることで生物炭素ポンプの効率に関与しているという興味深い仮説を提唱しています。また、私の所属する研究室でTara Oceansのデータを解析したところ、ウイルスの群集構造と生物炭素ポンプの効率との間に非常に強い相関があることが示されました。
また、この生物炭素ポンプの効率の鍵を握っていたのは、珪藻類やハプト藻類といった海洋の主要な植物プランクトンに感染するウイルスであるという結果が得られました(図2)。
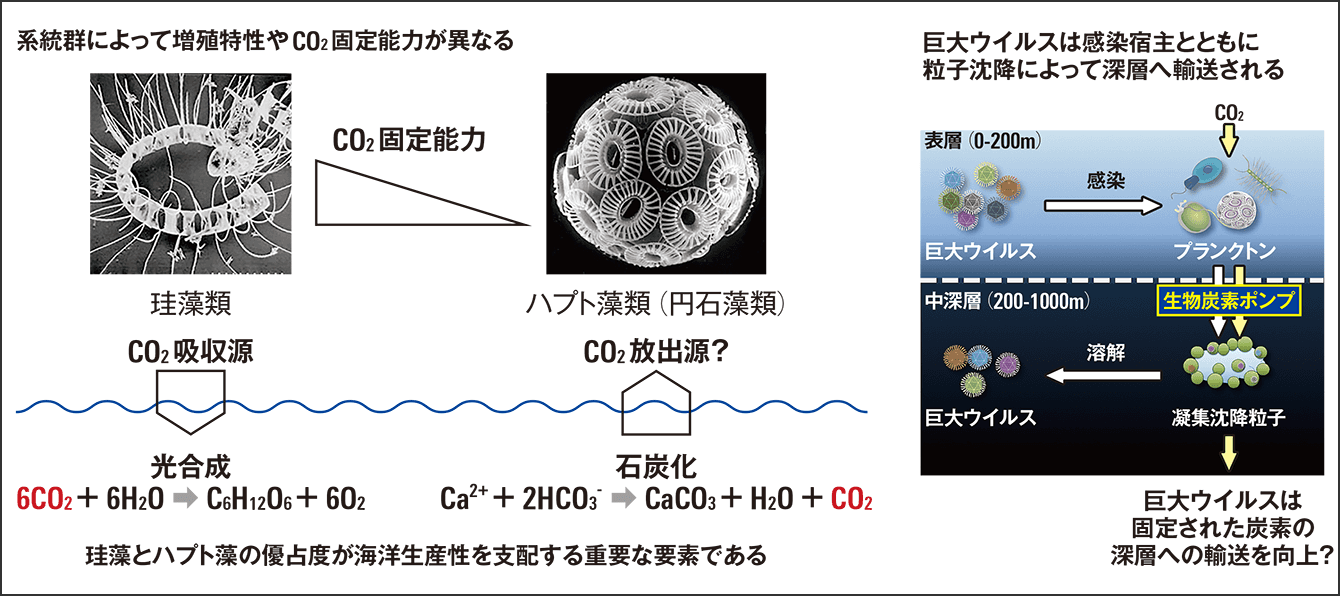
図2 珪藻類とハプト藻類:海洋における主要な植物プランクトン〈左〉主要な植物プランクトンの珪藻類とハプト藻類では、CO2固定能力が大きく異なる。〈右〉遠藤助教らの研究で、珪藻類やハプト藻類など主要な植物プランクトンに感染するウイルスが、CO2を海洋の中深層に固定する生物炭素ポンプの効率の鍵を握っていることがわかった。
珪藻類とハプト藻類は、海洋におけるCO2の固定能力について、対照的な働きをします。珪藻類は活発に光合成を行って、海水中のCO2分圧を下げ、大気からCO2が溶け込むのを促進します。一方で、ハプト藻類の石灰化を行う系統群(円石藻類)は石灰化の過程で海水中の炭酸塩緩衝作用を低下させ、大気からCO2が溶解しにくい環境をつくります。つまり、ハプト藻類が優占する海域は、光合成によるCO2の吸収能力が乏しい環境といえます。プランクトンの化石(微化石)を年代別に調査してみても、氷河期の寒い時期には珪藻類が優占していて、間氷期の暖かい時期にはハプト藻類の化石がたくさん出ています。
植物プランクトンは、巨大ウイルスの感染から防御するために、特定の化学物質を産生する可能性があります。例えば、藻類の代謝産物であるジメチルスルフォニオプロピオネート(DMSP)という有機硫黄化合物が細菌によって分解されると硫化ジメチル(DMS)が生じますが、この物質にはウイルス感染から宿主を保護する作用が報告されています。
興味深いことに、硫化ジメチルは水を引き込みやすい性質があり、これが大量に生成されれば大気に放出され、凝結核となって雲を発生させます。雲が形成されれば、太陽光を遮るので、地球を冷却する効果が唱えられています。特に、ハプト藻類はこのDMSPをよく生産することが知られています。
宿主のプランクトンが表層にたくさんいる生産性が高い海域は、それを殺せるウイルスもたくさんいます。ところが、ウイルスの効果が弱まり、生態系の循環が滞ると、「プランクトンの老化」ともいえる現象が起きて、放出する有機物の組成が変わってしまいます。その結果、雲が作られにくくなり、雨も降りにくくなる可能性があるというレポートがあります。
地球の気候システムはとても複雑なので、ウイルスの働きと温暖化とをシンプルにつなげるのは現時点では難しいかもしれません。しかし、植物プランクトンの生物量や群集組成が物質循環に与える影響については比較的よくわかっています。巨大ウイルスが植物プランクトン群集の制御因子であることはほぼ確実なので、その影響の規模が広域的かつ定量的に明らかになれば、将来起こる温暖化の規模や性質にウイルスがどの程度関与しているかという議論は可能だと思います。
今後、温暖化が進めば海洋表層はますます混ざりにくくなるため、深層から供給される栄養塩濃度は緩やかに減少していき、それに付随して植物プランクトンの生物量も減少すると予測されています。したがって、将来的な海洋のCO2吸収の機能を維持させる上でも、ウイルスと生物炭素ポンプとの関係解明が不可欠です。
そのために、私たちは、現在、世界で最も生物炭素ポンプの寄与が高い海域である太平洋の親潮域で、ウイルスによる生物炭素ポンプ駆動の実証を試みています。
(図版提供:遠藤 寿)