ミツバチの数が減っている。特に養蜂の主力であるセイヨウミツバチの減少が著しい。さまざまな原因が推察されるが、その中で直接的な影響を持つものに、ミツバチヘギイタダニが媒介する病気と天敵スズメバチがある。一方、日本の在来種であるニホンミツバチは、ダニに対する耐性があり、さらにスズメバチを撃退する熱殺蜂球と呼ばれる手段を有する。ニホンミツバチの遺伝子解析は、本種の保全、さらに、ミツバチの品種改良へと応用できる可能性があるという。
特集 ミツバチの世界 ニホンミツバチのゲノム解析がミツバチの減少抑制につながる!
構成/菊地武顕

東京都立大学理学部生命科学科特別研究員
若宮 健(わかみや・たけし)
2016年、京都産業大学総合生命科学部卒業。同大大学院生命科学研究科で修士課程、東北大学大学院生命科学研究科(生態発生適応科学専攻)で博士課程を修了。2021年、広島大学研究員。2022年、東京都立大学理学部生命科学科特任教員(特任研究員)。2023年4月から現職。
世界的にミツバチの数が減っているといわれています。特に養蜂に適しているセイヨウミツバチの減少が顕著です。その要因として、病原体の流行、気候変動、花資源の不足、天敵の侵入、家畜化による遺伝的劣化、農薬などが挙げられており、それらの要因がいろいろと複雑に絡んでいると考えられます。ニホンミツバチの減少数ははっきりとはしていません。ただニホンミツバチはセイヨウミツバチと異なる病原抵抗性を持つなどの特徴があり、注目を集めています。
ニホンミツバチの全ゲノム配列を比較
ニホンミツバチ(学名:Apis cerana japonica)はトウヨウミツバチ(A. cerana)の1亜種で、本州以南に生息する日本で唯一の野生種ミツバチです。本州、四国、九州、一部の離島と広く分布していますが、これまでの研究では、日本列島内で遺伝的に均一な集団であるとされてきました。しかし南北に長い日本列島は、気温差、標高差、降水量差、降雪量差など、ミツバチの生存に影響を与える条件を多く含みます。そのため、環境条件がニホンミツバチの遺伝的分化を促進し、独自の進化を起こしているのではないかという仮説を立てました。北限の青森県と南限の奄美大島との間には幅広い分布レンジがあるので、それぞれに異なった適応集団がいるのではないかと考えたわけです。
そこで私はこれまでの研究とは異なり、最新の全ゲノム解析を試みました。その結果、ニホンミツバチは大きく3つの地域で異なる遺伝的特徴があることが分かりました。
これまで行われてきた研究は、形態学的アプローチと遺伝学的アプローチによる地域差の比較です。
形態学的アプローチでは、クラスター解析という異なるものが交ざり合っている集団の中から似たもの同士を集めてグループ(クラスター)分けする分類方法によって、翅脈の長さを解析するのが一般的なのですが、地域性は確認されていませんでした。また岩手県、鹿児島県、奄美大島の3カ所での体サイズを比較した先行研究では、本土–奄美大島間では有意差が確認されましたが、岩手県と鹿児島県という離れた地域で有意差が確認されませんでした。
また、遺伝学的アプローチでは、ミトコンドリアDNAのDループ領域などで比較が行われてきました。ミトコンドリアという細胞内小器官に存在するDNAの中にあるDループと呼ばれる領域は、塩基置換が頻繁に見られ、他の領域と比較し進化速度が速いといわれますが、この比較でも明瞭な地域差は出ていなかったのです。
先行研究では、ゲノム上の多型を数カ所しか比較できず、特定の形質や変異しか比べられていませんでした。これをゲノムスケールにアップデートすることで、数百万カ所の多型を見ることができますし、特定の形質や変異だけに目を向けるのではなく全変異を調べることで変異の意味づけも可能となります。
実際、日本列島における昆虫の地域適応について、これまでもゲノムスケールでの研究がいくつかなされています。例えばゲンジボタルは、九州から東北の地域にかけていくつかの集団に分かれ、さらに、西日本と東日本とでは点滅のタイプも異なります。
3つの地域で異なる遺伝的特徴
私たちの研究グループは青森県から鹿児島県まで日本各地の105個体のニホンミツバチの全ゲノム配列を解析しました(図1)。全ゲノムの中から、DNA配列に違いがある一塩基多型(SNP)を約127万カ所見つけ、それを詳細に比較。その情報を使って遺伝構造の推定をしたのです。
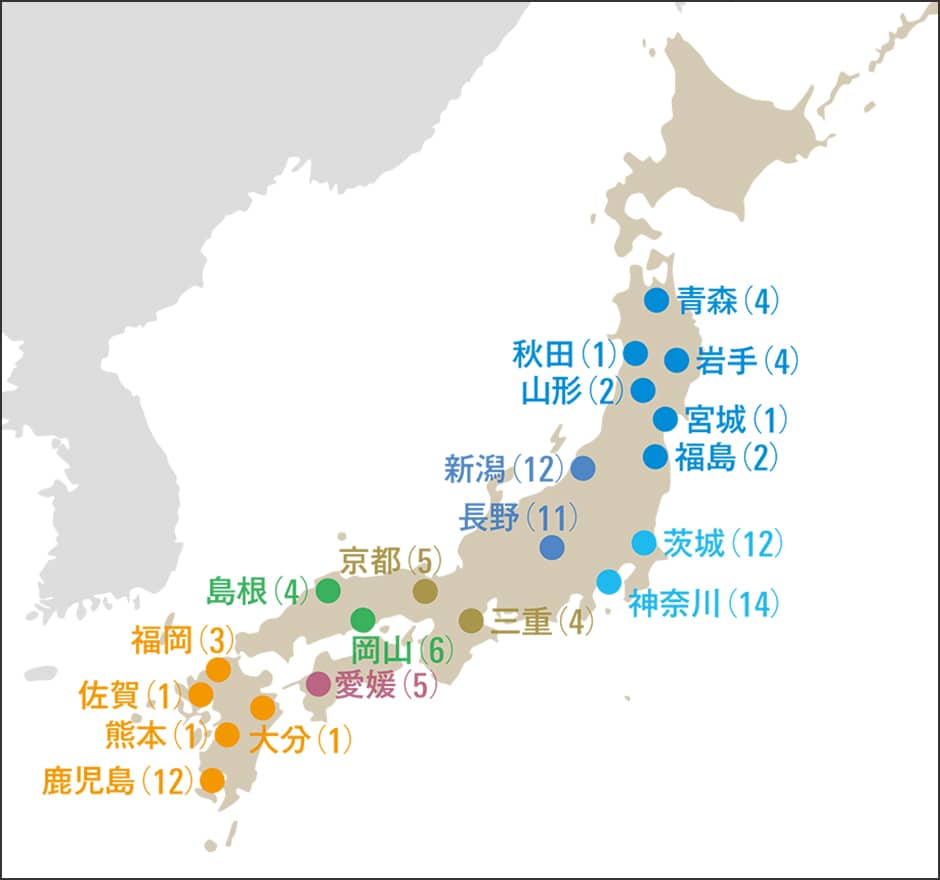
図1 ゲノム解析をしたニホンミツバチの分布農業・食品産業技術総合研究機構(農研機構)と養蜂家の協力を得て、青森県から鹿児島県まで日本全域から105個体のニホンミツバチのゲノムを解析。大部分の飼育群は各地域の自然群に由来する。
まずアドミクスチャー(ADMIXTURE)解析といって、集団をいくつかの近似した部分ごとに分ける解析を行い、遺伝的組成を可視化してみました。その際に、中国のトウヨウミツバチとの比較も行ったところ、ニホンミツバチとの違いは明らかでした。日本と大陸との分断は、だいたい10万年前といわれています。それ以降の年代でニホンミツバチの遺伝構造が最終的に固定されてきたといえるでしょう。
この解析で、日本列島の集団は大きく北部(東北・関東・中部地方)、中央部(中国地方)、南部(九州地方)の3つの地域に区別できることが明らかになりました。北部は青色が、近畿と四国は複数色が交じり合い、中央部は緑、南部の九州はオレンジが大きく占めていることが分かります(図2)。大規模ゲノム情報で、このような多様性の存在が明らかになったのです。
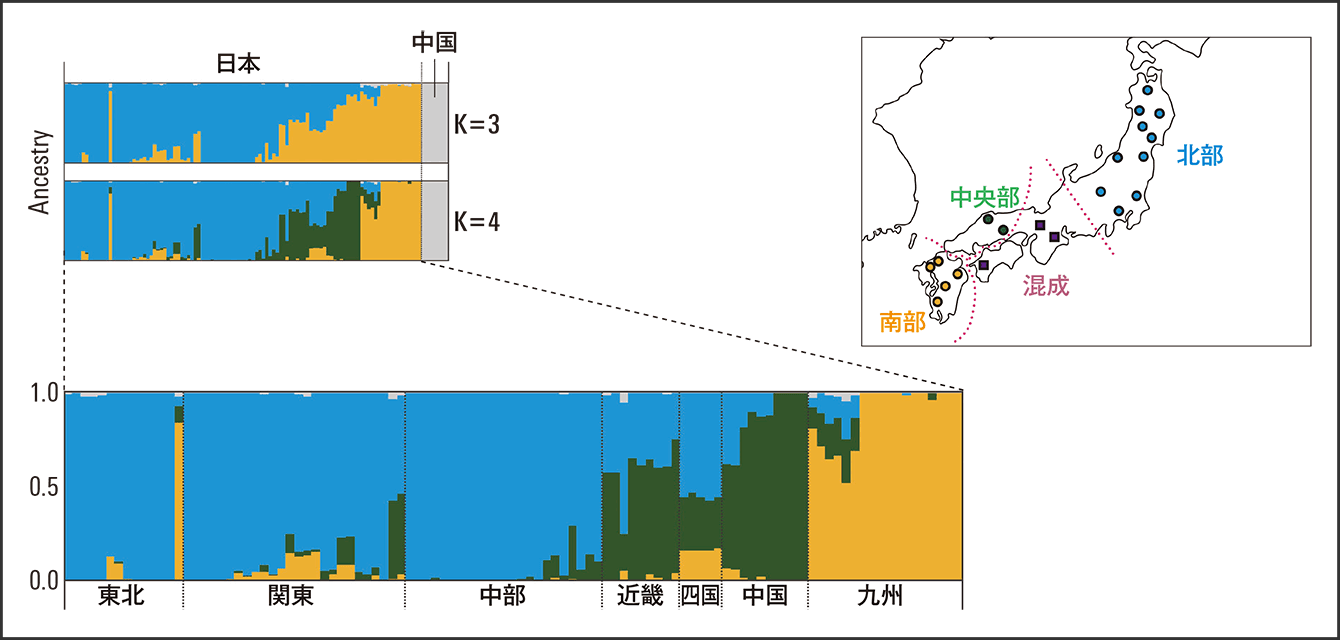
図2 日本列島で確認されたニホンミツバチの特徴的な遺伝構造左上図のように、中国のトウヨウミツバチとニホンミツバチとの相違は明らかだった。東北・関東・中部地方の北部、中国地方の中央部、九州地方の南部の3地域ではっきりとした違いが出た。近畿、四国地方では混成している。
中には例外的な個体もありました。例えば北部の福島県のミツバチなのに、南部と同様の遺伝的組成を持つ個体がいたのです。そこでこのハチの“出自”について調査をしたところ、実は熊本県から福島県に導入されたものだということが分かりました。他の例外も、他地域からの人為的移入が疑われるものが大半でした。
これは、異なる地域間で個体を移動させると、移動した蜂群が新たな環境に適応できない可能性があることを示唆するものです。トウヨウミツバチは趣味で養蜂する人が多く、そうした愛好家同士で交流し、ハチを送り合ったり、売買することもあるようです。本来の適応状態と合っていない地域にハチを送り込んでいる状況も、今回の調査で検出できました。
3地域のそれぞれで、自然選択を受けた遺伝領域はどこかについても、探索してみました。集団ベースの分化ならびに環境変数との相関について調べてみたのです。
まず集団ベースの分化では、北と中央部では似ているが南だけ分かれている領域、あるいは中央部と南は似ているが北だけが分かれている領域など、地域特異的な選択領域を検出してみました。それを見つけ出すことで、どういう遺伝領域がその地域への適応に関与したかを知ることができるからです。
その結果、南部の集団で特異的に分化した遺伝領域に、シェーカー(Shaker:potassium voltage-gated channel protein Shaker isoform X3)がありました。これはショウジョウバエの異常行動や寿命、睡眠などに関連するといわれる遺伝子です(図3)。中央部の集団で特異的な分化をした遺伝領域には、GDPGP1、CBSなど9遺伝子がありました。GDPGP1はグリコーゲン量を制御し、CBSはメチオニン代謝に重要で、エネルギー代謝と関連があります。北部の集団では、NPC2、HYPKなど6遺伝子が特異的に分化していました。NPC2は病原体への感染時に発現量が変動することが報告されていて、HYPKはアポトーシス(細胞死)と関連があり、免疫機能と関わっている可能性があるでしょう。
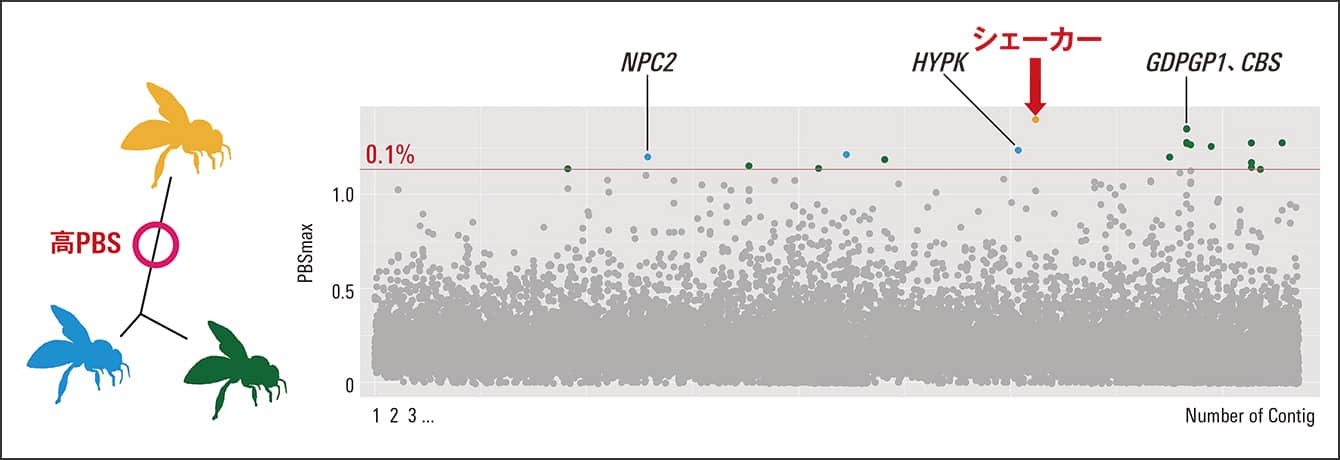
図3 南部集団で特異的な分化の遺伝領域(シェーカー)PBS(Population Branch Statisitcs)は遺伝的分化度を指標とした統計量の1つで、全ゲノムを一定の範囲ごとに区切ってスコアを算出した。上位0.1%の外れ値ウインドウの領域から計25遺伝子が検出された。南部集団ではシェーカーの周辺領域で特異的な分化が認められた。
続いて、気温、積雪量、降水量、日照時間といった環境の違いと遺伝子の相関も調べました(図4)。例えば、採集地点の平均気温が上がるにつれて、遺伝子の型がグラデーションを描くように変化するパターンを検出します。こちらの環境変数との相関をベースとした解析では約70個の候補遺伝子を検出することができましたが、集団ベースの分化と関わる25個の遺伝子との共通項は存在しませんでした。このことは、ニホンミツバチは各地域の特異的な環境に適応しており、今後、気温が上昇したからといって単純に北へ移動するのが困難なことを示唆しています。
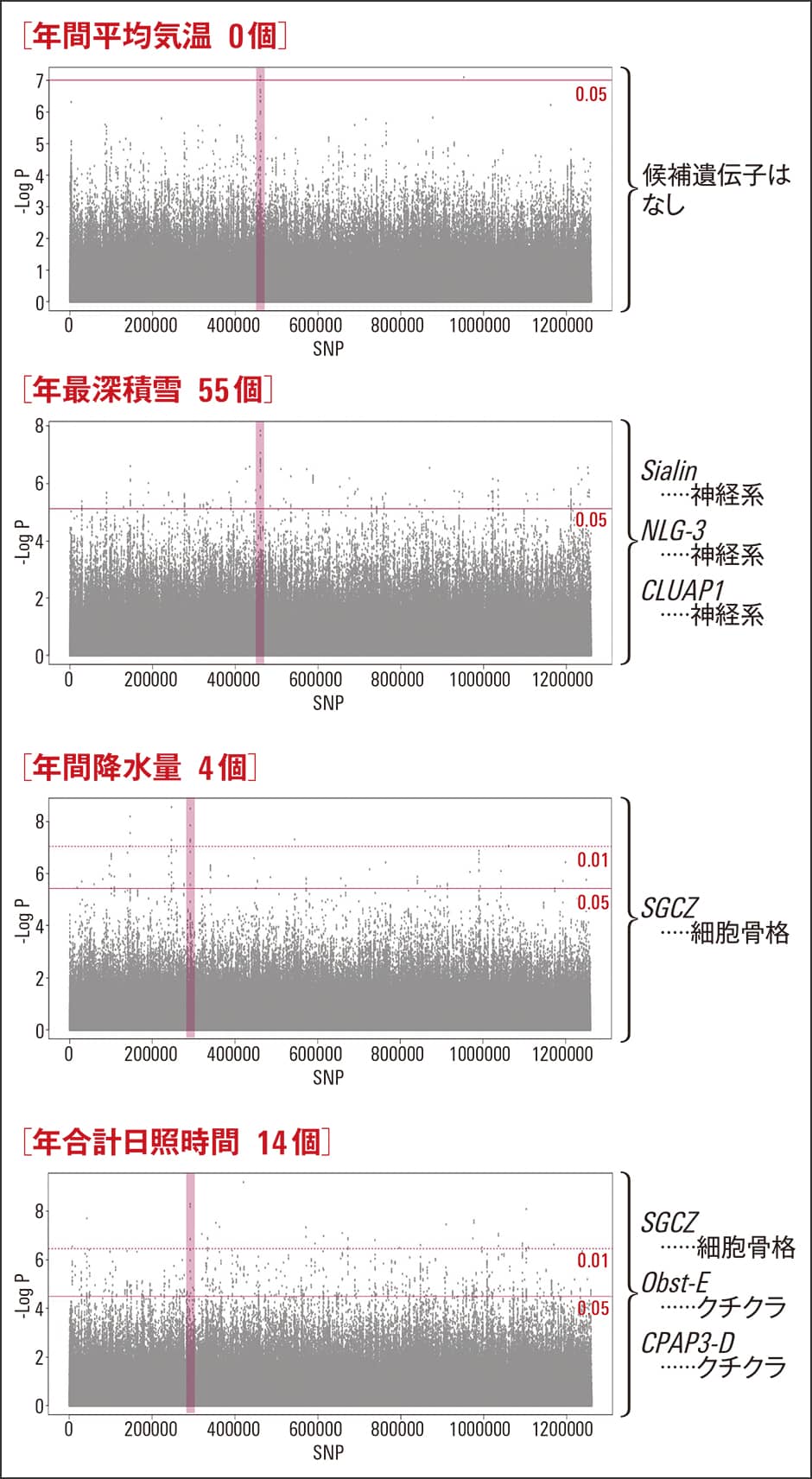
図4 環境の違いと遺伝子の相関関係環境変数と相関する候補遺伝子の検出を試みた。意外なことに、年間平均気温と相関する候補遺伝子は特定できなかった。一方で積雪量については、Sialin、NLG-3、CLUAP1といった神経システムと関連する候補遺伝子を複数検出した。
ミツバチ進化ゲノミクス研究の今後の展望
ニホンミツバチの遺伝的分化と局所適応について、ようやくここまで分かってきたところです。今後、3つの地域集団のどこに由来するのかを判別する簡易的なシステムを開発することで、適切な移動制限や地域適性の診断などが可能になると期待されます。また、局所適応に関わる候補遺伝子が、具体的にどのような環境で、どのように適応に関与しているのか(例えば、とある環境下での活動性、花の好み、病気への耐性など)を調べることで、ニホンミツバチの保全および応用利用の両面での貢献が可能になると考えられます。
養蜂種であるセイヨウミツバチの病気の代表例として、ミツバチヘギイタダニが原因のバロア症があります。このダニが幼虫やサナギに寄生し体液を吸うことで、発育障害を起こすのです。またミツバチヘギイタダニは、チヂレバネウイルスなど少なくとも5種類のウイルスの宿主でもあるのです。チヂレバネウイルスに感染した幼虫は、羽化時に翅が正常に伸展せずに死亡します。ニホンミツバチを含むトウヨウミツバチが病気に強いといわれる理由の一つは、ミツバチヘギイタダニに対する耐性があるからです。
もともとミツバチヘギイタダニは、トウヨウミツバチの寄生虫でした。それが、養蜂のためにアジア各国に導入されたセイヨウミツバチに移動したのです。ニホンミツバチとセイヨウミツバチは、採餌のために花のある場所で競合しますし、ニホンミツバチがセイヨウミツバチの巣に入り蜜を盗んだりすることも観察されています。そのように2種のミツバチが接したときに、ミツバチヘギイタダニも移動可能です。その後、ミツバチの国際的な輸出入の際にミツバチヘギイタダニも随伴し、世界各地に広がったと考えられています。どう移動していったかについても、遺伝子をたどって明らかにされています。英国サルフォード大学のスティーヴン・J・マーティン教授らの研究グループが、病原体遺伝子解析についての論文を発表しています。
セイヨウミツバチの減少要因として、天敵の存在も挙げられています。その一つがスズメバチです。ニホンミツバチは、巣にやって来たスズメバチに熱殺蜂球と呼ばれる防衛手段を用いて抵抗します。1匹のスズメバチを数百匹のニホンミツバチが包み込むように囲んで、一斉に飛翔筋を動かすことで熱を発生させ、蜂球内の温度を45~47℃まで上昇させます。20~40分間にわたって熱を浴びることで、スズメバチは熱死します。スズメバチとニホンミツバチとでは致死温度に3℃ほど差があり、ニホンミツバチはこの温度差によってスズメバチを撃退するのです。
セイヨウミツバチも似たような行動をするのですが、熱殺蜂球をうまくつくる前に襲われて全滅してしまいます。スズメバチ類はアジアを中心に生息しています。ずっとアジアで生息してきたトウヨウミツバチと、スズメバチの少なかった地域で生息してきたセイヨウミツバチの違いがこうした性質に表れているのでしょう。
セイヨウミツバチのゲノムは、ニホンミツバチに先行し、すでに詳細な解析が進んでいます。今後、ゲノム解析でニホンミツバチとセイヨウミツバチの特徴を比較し、例えばダニに強い耐性を持つ遺伝子を特定することができれば、品種改良をしてダニ由来の病気に強いセイヨウミツバチが実現できるかもしれません。また、熱殺蜂球の形成に重要な遺伝子を特定できれば、スズメバチに強い性質をセイヨウミツバチに持たせられる可能性があります。
人間はミツバチから大きな恩恵を受けています。ハチミツやローヤルゼリーといった産物を食べるだけではありません。大きな恩恵は、花粉交配です。ミツバチのおかげで、世界の食料生産の3分の1が支えられているといわれています。これからの世界的人口増で食料不足が懸念されている中、ミツバチの減少は切実な問題となるのではないでしょうか。そのためにもゲノム解析などの先端技術を用いてニホンミツバチのことをより詳しく知ることが大事だと感じています。
(図版提供:若宮 健)